真菌毒素是由曲霉菌属、镰刀菌属或青霉菌属在生长繁殖过程中产生的有毒次级代谢产物,能够污染几乎所有种类的食用、饲用农产品以及中草药等,严重危害人畜健康。据统计,全球25%的粮食作物受到真菌毒素的污染,造成的经济损失达数千亿美元/年;我国每年在生产、储存、运输、销售过程中受到真菌毒素污染的粮食有3100多万t。因此,如何有效控制和消除食品、饲料中的各类真菌毒素,成为食品、饲料行业亟待解决的问题。
据统计,目前对玉米、大豆、小麦以及饲料污染**严重的真菌毒素主要有黄曲霉毒素、赭曲霉毒素、玉米赤霉烯酮、伏马毒素和呕吐毒素,这几种毒素的控制和降解成为各国政府关注的焦点。
1.1 黄曲霉毒素
黄曲霉毒素(aflatoxin,AF)是由曲霉产生的一种二呋喃氧杂萘邻酮的衍生物,在紫外光下可发出荧光,根据荧光颜色的差异,分为黄曲霉毒素B(AFB 蓝色荧光)、黄曲霉毒素G(AFG 绿色荧光)和黄曲霉毒素M(AFM 蓝紫色荧光)等 。黄曲霉毒素是目前发现的化学致癌物中**强的物质之一,主要通过损害肝脏导致肝癌。此外,黄曲霉毒素还可诱发直肠癌、乳腺癌和骨癌等,其一般存在于玉米、花生、棉籽、稻谷以及大豆等多种农作物中,以玉米和花生中的污染**为严重。而在黄曲霉毒素的几种构型中,AFB1的毒性**强 ,AFM1、AFG1、AFB2和AFG2 的毒性相对较弱。目前,全球已有100 多个国家和地区针对食品和饲料中黄曲霉毒素的限量制定了相关标准。美国标准规定AFM1 在牛奶和饲料中的含量分别不能超过0.5 和300 μg/kg,而AFB1、AFB2、AFG1 和AFG2 在食品中的总量不超过20 μg/kg ;欧盟标准规定乳制品中AFM1 含量不能超过0.05 μg/kg, 食用花生仁中AFB1、AFB2、AFG1 和AFG2 总含量不超过4 μg/kg 。我国也对黄曲霉毒素在各类农产品和食品中的含量有着严格的标准,根据GB 2761—2011 的有关规定:AFB 在玉米中的限量为20 μg/kg,在大米和稻谷中为10μg/kg,其他粮食作物中则为5 μg/kg;对于婴儿食品而言,AFB 和AFM 的限量都仅为0.5 μg/kg 。
1.2 赭曲霉毒素
赭曲霉毒素(ochratoxin,OT)是由曲霉或青霉产生的有毒次级代谢产物,存在A(OTA)、B(OTB)、C(OTC)和α(OTα)等衍生结构,其中OTA 衍生物毒性**强,对农作物的污染**为严重 。赭曲霉毒素主要对肾脏和肝脏产生损伤,具有致癌性、免疫毒性和致畸性等潜在危害,其主要存在于小麦、玉米和花生等农作物中。目前,多个国家或地区对食品和饲料中赭曲霉毒素的限量制定了相关标准:欧盟规定饲料用谷物中OTA 的含量不能超过0.25mg/kg,猪补充饲料和配合饲料中OTA 的含量不超过0.05 mg/kg ,我国GB 2761—2011 对OTA 在各类谷物及豆制品中的限量为5 μg/kg 。
1.3 玉米赤霉烯酮
玉米赤霉烯酮(zearalenone,ZEN) 是由镰刀菌属产生的次级代谢产物,主要存在于玉米、小麦、大麦和大米等谷物中,具有较强的耐热性,在110 ℃条件下需要处理1 h 才能被完全破坏。此外,玉米赤霉烯酮作为雌激素的类似物,能够作用于动物的生殖系统,导致其繁殖机能受到损害,甚至引发死亡。目前,各国对玉米赤霉烯酮的限量尚未达成统一标准,我国对ZEN 的限量有着严格的规定。根据GB 2761—2011 的标准,ZEN 在各类农产品中的限量均为60 μg/kg 。欧盟对于ZEN 在玉米内的限量为3 000 μg/kg,在其他谷物中为2 000 μg/kg,而在猪饲料中的限量很低,仅为250 μg/kg。
1.4 伏马毒素
伏马毒素(fumonisins,FB) 是由串珠镰刀菌产生的一种水溶性代谢产物,已发现的伏马毒素有FA1、FA2、FB1 和FB2 等共11 种,其中FB1 **为常见,主要污染玉米及玉米制品。伏马毒素能够损害肝肾功能,且是一种致癌物,与食道癌的高发有一定的关系。与玉米赤霉烯酮类似,目前国际上对食品和饲料中伏马毒素的限量及检测方法仍无统一标准:欧盟针对饲料原料、玉米及其产品中伏马毒素(FB1 +FB2)的指南限量为60 mg/kg,补充饲料和配合饲料中伏马毒素(FB1 +FB2 )的指南限量为5mg/kg;美国FDA 规定食用家禽饲料(玉米及玉米副产品)中伏马毒素(FB1 +FB2 +FB3)的指南限量为100 mg/kg,猪饲料(玉米及玉米副产品)中伏马毒素(FB1 +FB2 +FB3 ) 的指南限量为20 mg/kg;此外,中国GB 2761—2011 并没有提及关于伏马毒素的限量标准。
1.5 呕吐毒素
呕吐毒素(deoxynivalenol,DON)是一种由镰刀菌产生的单端孢霉烯族化合物,主要存在于小麦、大麦和玉米等谷物籽实中,耐热性较强,121 ℃高压加热25 min 仅有少量破坏。呕吐毒素对哺乳动物具有较强的毒性,能够引起人和动物消化系统疾病和厌食症,猪对呕吐毒素**为敏感,家禽次之,反刍动物耐受能力**强 。近年来,各国纷纷意识到呕吐毒素的危害,制定了相应的标准,***农副产品中呕吐毒素的含量:根据美国FDA 标准,饲料用谷物及其副产品(除玉米外)中呕吐毒素的限量≤1mg/kg;欧盟规定玉米及其副产品允许≤1.75mg/kg的呕吐毒素残留;中国卫生部对农副产品中残留的呕吐毒素规定为不得高于1mg/kg。
由于产生真菌毒素的微生物所偏好的生长环境不同,真菌毒素在全球的分布具有一定的地域性。热带和***带是曲霉生长的**佳环境,而镰刀菌**适宜在北美、欧洲和亚洲的气候条件下生长。在炎热、气候潮湿的热带地区,黄曲霉毒素的污染比较严重。玉米赤霉烯酮、赭曲霉毒素和伏马毒素主要在温带区域,如中国、欧洲及北美洲等地区。2016年,百奥明公司针对81个国家16511个样品的检测发现,非洲作物中黄曲霉毒素的污染**为严重,检出率为60%;北美玉米中伏马毒素的检出率高达66%,南美作物中主要的毒素为玉米赤霉烯酮和呕吐毒素;中东作物中存在的主要毒素为呕吐毒素和伏马毒素;欧洲作物中的主要毒素类型为呕吐毒素,验证了真菌毒素的区域分布特性。但近年来随着国际贸易进程的加速,各类毒素的分布正逐渐呈现全球化的发展趋势。
对真菌毒素超标的农产品,**简单的处理方法是丢弃或销毁,但是,这将造成巨大的浪费和经济损失,同时也会产生严重的环境污染。因此,迫切需要开发安全、**、低成本的真菌毒素脱除方法。常用的真菌毒素脱除方法有物理法、化学法和生物法。
化学法是采用酸、碱、氧化剂、醛或亚硫酸气体以改变真菌毒素的结构,该方法虽然能够有效脱除毒素,但可能会对食品的营养价值和风味产生影响,且存在化学物残留的安全隐患。物理脱毒法是采用吸附剂将毒素进行脱除,但该方法稳定性差,且目前商品化的吸附剂对霉菌毒素的脱除效果仍有待加强。
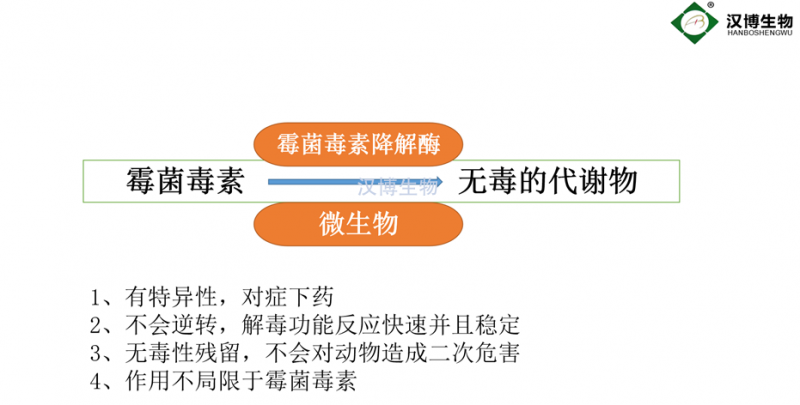
生物法脱毒包括微生物法和生物酶法,前者是利用微生物对毒素的吸附或代谢能力,实现毒素的脱除,成本相对低廉,但并不适用于所有领域,如食品加工过程等。生物酶法脱毒是从微生物中发掘降解毒素的关键基因,利用基因技术构建生物酶的**表达工程菌,分离获得纯酶以进行食品和饲料中真菌毒素的脱除。与微生物脱毒法相比,酶法脱毒具有更好的重复性、均一性和操作简单等特点,受到了广泛关注。
研究者针对黄曲霉毒素、赭曲霉毒素、玉米赤霉烯酮、伏马毒素和呕吐毒素均开发了相应的降解酶(表1),下文将具体阐述。
3.1 黄曲霉毒素的酶法降解
黄曲霉毒素的生物降解酶主要为氧化还原酶类。1998 年,Liu 等发现在真菌Armillariella tabescens中存着可降解黄曲霉毒素的复合多酶体系,并在随后的研究中分离获得起降解作用的关键酶黄曲霉毒素氧化酶(AFO)。该酶主要作用于AFB1 的二呋喃环,从而破坏黄曲霉毒素。Doyle 等报道寄生曲霉能够产生降解黄曲霉毒素的乳过氧化物酶,并且黄曲霉毒素的降解率与酶含量成正相关性。Yehia从白腐真菌Phanerochaete ostreatus中分离出一种能降解AFB1 的锰过氧化物酶,其对AFB1 的脱毒效率依赖于酶的浓度以及酶反应的时间,在**优条件下,1.5 U/ mL 的酶能够在48 h 内降解90%的AFB1。此外,漆酶亦被发现具有降解黄曲霉毒素的功能, Alberts 等 研究发现来自于白腐真菌Phanerochaete ostreatus的漆酶能够降解黄曲霉毒素,118 U/L 的漆酶对黄曲霉毒素的降解率为55%。Loi 等亦从P.pulmonarius和P.eryngii 中分离获得了漆酶,将其用于AFB1 和AFM1 的降解,发现仅存在漆酶的情况下,上述毒素的降解效率较低,而当加入10 mmol/ L 氧化还原介质后,反应72 h,对AFB1 的降解效率可从23%提高至90%,对AFM1 的降解效率则可提高至100%。2010 年,Novozymes 公司对氧化还原介质介导的漆酶降解黄曲霉毒素进行了专利申请,该专利中漆酶来源于Streptomyces coelicolor,丁香酸甲酯作为氧化还原介质,在此条件下,反应24 h, 漆酶对黄曲霉毒素的降解率达100%。2017 年,Xu 等从43 株菌株中筛选获得了Bacillus shackletonii L7,在37 ℃反应72 h,Bacillus shackletonii L7 对AFB1、AFB2 和AFM1 的降解率分别为92.1%、84.1%和90.4%。在此基础上,Xu 等进一步分离获得了能够降解黄曲霉毒素的新酶(芽孢杆菌黄曲霉降解酶),该酶蛋白的分子量为2.2×104,**适反应温度和pH 分别为70 ℃和8.0。
3.2 赭曲霉毒素的酶法降解
降解赭曲霉毒素的生物酶主要为羧肽酶,具体为羧肽酶A 和羧肽酶Y。其中,羧肽酶A 是**早被发现的具有赭曲霉毒素降解功能的生物酶,来源于牛胰腺中,对赭曲霉毒素的亲和性相对较高,25 ℃条件下Km值为1.5×10-4 mol/ L。羧肽酶Y来源于酿酒酵母,其对赭曲霉毒素的降解能力相对较弱,5 d 仅能降解52%的赭曲霉毒素。此外,脂肪酶、蛋白酶和酰胺酶等亦被发现具有降解赭曲霉毒素的功能。Abrunhosa 等从黑曲霉中分离获得了可以降解赭曲霉毒素的纯酶,该酶的**适反应温度和pH 分别为37 ℃和7.5,在此条件下,其对赭曲霉毒素的降解活性高于羧肽酶A。Stander 等通过对系列水解酶的筛选,发现来源于黑曲霉的脂肪酶对赭曲霉毒素具有较高的降解能力,其对赭曲霉毒素的比活力达2.32 U/ mg。Abrunhosa 等的研究发现,一些商品化的酶亦具有降解赭曲霉毒素的能力,在pH7.5 条件下反应25 h,蛋白酶A 对赭曲霉毒素的降解率为87.3%,胰酶对赭曲霉毒素的降解率为43.4%。Yu 等的专利中报道酰胺酶能够降解赭曲霉毒素,当用160 ng/ mL 的酰胺酶降解50μg/ mL 的赭曲霉毒素,降解率可达83%。
3.3 伏马毒素的酶法降解
早在1996 年,Duvick 等已从E.spinifera 中分离获得可降解伏马毒素的基因,并将其克隆至玉米等农作物中,在相关酯酶、胺氧化酶及其他酶的作用下,伏马毒素被逐渐水解、氧化。2009 年,Heinl等研究发现在Sphingopyxis sp.MTA144 的同一个基因簇上存在编码羧酸酯酶和转氨酶的2个基因,在这2个酶的作用下,FB1 经二步酶促反应被降解:在羧酸酯酶的作用下FB1 被降解为HFB1,在转氨酶的作用下HFB1 发生转氨反应,并**终转化生成2-酮基-HFB1。Hartinger 等将Sphingopyxis sp.MTA144 中编码转氨酶的基因FumI 在大肠杆菌中进行了表达,并对其酶学性质进行了研究,发现该酶的**适温度为35 ℃,**适pH 为8.5。2011 年,Heinl 等从Sphingopyxis sp.ATCC 55552 克隆获得了新的转氨酶基因,并将其在大肠杆菌中进行了表达,发现该酶亦具有降解HFB1的能力。百奥明研究中心从Sphingopyxis sp.MTA144 中分离获得了酯酶,并开发出活性成分为纯酶的FUMzyme® ,这种酶制剂在动物的胃肠道内能够有效降解伏马毒素为毒性显著降低的化合物。
3.4 玉米赤霉烯酮的酶法降解
能够降解玉米赤霉烯酮的生物酶主要有三类:漆酶、内酯水解酶和过氧化物酶。其中来源于 Trametes versicolor 的漆酶,反应4 h 对玉米赤霉烯酮的降解率为81.7%。而在2009 年,Novozyme 公司的Viksoe-Nielsen 等发现在氧化还原介质的存在下,来源于Streptomyces coelicolor的漆酶除了能降解黄曲霉毒素外,还能降解玉米赤霉烯酮,所采用的氧化还原介质可以为丁香酸甲酯,37 ℃条件下反应24 h,玉米赤霉烯酮可被完全降解。目前,对玉米赤霉烯酮降解酶**为关注的酶为内酯水解酶,编码基因为zdh101,是Takahashi-Ando 等2002 年从来源于Clonostachys rosea IFO 7063 中分离获得,并将其在酿酒酵母中实现异源表达,脱毒实验表明其能够在37 ℃条件下反应8 h 或28 ℃条件下反应48 h,实现2 μg/mL 玉米赤霉烯酮的完全降解。唐语谦等发现该酶主要通过作用于玉米赤霉烯酮的内脂键实现毒素的降解,但由于可逆反应的存在,该反应并不能完全降解玉米赤霉烯酮,降解产物为β-ZOL,仍具有一定雌激素毒性,但毒性相对较低。此外,Yu 等研究发现来源于Acinetobacter sp.SM04 的过氧化物酶(Prx)能催化H2O2 氧化降解玉米赤霉烯酮。在0.09%H2O2 存在的条件下,30℃反应6 h 能降解玉米样品中90%的玉米赤霉烯酮。目前,该酶具体的作用机制仍在研究中。
3.5 呕吐毒素的酶法降解
呕吐毒素降解主要有3 种途径,分别为糖基化、氧化和乙酰化,这3 种反应又有其对应的酶进行催化。Poppenberger 等于2003 年发现了一种典型的呕吐毒素糖基化酶——Arabidopsis thaliana 中的UDP-糖基转移酶,能通过催化葡萄糖从UDP 葡萄糖转移到呕吐毒素的C3 位的羟基上形成3-O-吡喃葡萄糖基-4-DON。2010 年,Schweiger 等将A.thaliana 中编码UDP 糖基转移酶的基因在拟南芥体内进行表达,培养出具有DON 抗性的农作物。Ito 等于2013 年在Sphingomonas sp.KSM1 中发现的P450 细胞色素系统能够将呕吐毒素氧化为16羟基脱氧雪腐镰刀菌烯醇(16HDNO),从而使其失去毒性。进一步通过相关基因的胞外重组和表达,发现了这一系统主要由Cyt450 的编码基因ddna及其对应的内源性还原物组成,其催化效率为6.4mmol/(L·s),对于质量浓度约为100 μg/mL 的呕吐毒素,这一系统能在3 d 内达到将近100%的降解率。另一种降解呕吐毒素的方法是将其3 号位乙酰化。Kimura 等于1997 年发现了一种能催化呕吐毒素乙酰化的酶——单端孢甲-3-O-乙酰转移酶并确定了其对应的编码基因,是一种从大肠杆菌中提取、由Tri101基因编码的乙酰化酶,它能够催化呕吐毒素3 号位乙酰化以达到解毒的目的。
3.6 不同真菌毒素及其相应的降解方法
上文中主要阐述了各类真菌毒素的生物降解方法,这些方法中涉及了相关的微生物、酶及反应,现将上述内容总结,见表1。
目前虽然已有能够降解真菌毒素的相关酶的报道,但已实现商业化应用的仅为能降解伏马毒素的FUMzyme®,大部分酶因为降解效率低、稳定性差等原因处于实验室研究阶段,针对真菌毒素降解酶的挖掘需进一步的深入。
目前所发现的真菌毒素降解酶,主要是基于色谱分离法获得。研究者在筛选获得可降解真菌毒素微生物的基础上,采用色谱法分别对微生物的胞外酶和胞内酶进行分离,将分离的各个组分分别进行真菌毒素降解实验,逐步确定起降解作用的关键酶。Zhao 等采用乙醇沉淀、色谱分离的方法从Myxococcus fulvus ANSM068 的胞外酶中分离获得了可降解黄曲霉毒素的酶,经测定该酶的分子量为3.2×104,**佳反应温度为35 ℃,**佳反应pH 为6.0,对AFG1 的降解率可达96. 96%,对AFM1 的降解率可达95.8%。Abrunhosa 等采用丙酮沉淀和色谱分离法从黑曲霉中分离获得可降解赭曲霉毒素的新酶,该酶的**适pH 为7. 5,**适反应温度为37 ℃,对赭曲霉毒素的**大反应速率Vmax 为0. 44μmol/(L·min)。虽然基于色谱分离技术能够筛选获得具有降解真菌毒素功能的新酶,但蛋白分离过程复杂,回收率低,需进一步对分离所得的蛋白进行鉴定,费时费力。虽然色谱分离法能够获得毒素降解相关酶,但由于微生物代谢过程非常复杂,目标蛋白获取的难度较大,过去10 年中被报道的具有脱除真菌毒素功能的新酶仅有7 个,需要开发更**的毒素降解酶发掘方法。
随着测序技术的发展和成本的不断降低,利用高通量测序技术开展真菌毒素降解菌基因组信息的检测已逐渐成为一种主流方式。2012 年,Zhou等发现菌株Devosia 具有降解呕吐毒素的能力,此时尚未见任何关于Devosia菌株基因组信息的报道,但截至目前,上传至GenBank 中的Devosia全基因序列达12 个。不断丰富的基因信息库以及组学技术的发展为真菌毒素降解酶的发掘提供了新的策略。Liuzzi 等在前期研究过程中筛选获得了一株能够降解赭曲霉毒素的不动杆菌ITEM 17016,为筛选获得降解赭曲霉毒素的关键基因,分析了ITEM17016 在含/不含毒素培养基中生长时的基因表达差异,发现有6 个肽酶在含毒素的培养基中表达上调,进一步将编码羧肽酶PJ 1540 的基因在大肠杆菌中进行异源表达,从而发掘了降解赭曲霉毒素的新酶。随着宏基因组技术、转录组学技术等的不断发展和成熟,预计未来越来越多具有降解毒素功能的酶将被挖掘。
目前针对酶法降解真菌毒素的研究主要集中于新酶的发掘以及酶学性质的研究,但天然酶在食品、饲料中的应用可能由于环境条件的***,无法获得理想的效果,往往需要采用蛋白质工程对酶进行改造,如增加酶的活性,pH、温度稳定性等,生物信息学的发展为脱毒酶结构的解析及改造提供了有力的技术支撑。Dobritzsch 等从黑曲霉中发掘了一种可以降解赭曲霉毒素新酶,该酶具有较高的热稳定性,**适反应温度为66 ℃,**适反应pH 为6,且比羧肽酶A 和羧肽酶Y 具有更高的降解性能;在此基础上,进一步对该酶进行了纯化、结晶和结构解析,发现该酶属于酰胺水解酶家族,具有双金属中心催化位点。通过对粉红粘帚菌的内酯水解酶ZHD101 的纯化和晶体培养,Peng 等解析了ZHD101 的分子结构,发现ZHD101 属于α/β-水解酶家族,分子结构由催化核心结构域和α-螺旋帽子结构域组成,底物结合在二者之间的深口袋中,靠近催化三联体为Ser102-His242-Glu126,酶与底物的作用是通过氢键和非极性键的互作完成。玉米赤霉烯醇为玉米赤霉烯酮的衍生物,其中α-玉米赤霉烯醇的雌激素较玉米赤霉烯酮具有更高的毒性,而ZHD101 能够同时降解玉米赤霉烯酮和α-玉米赤霉烯醇,但对后者的降解效率仅为前者的40%,为提高ZHD101 对α-玉米赤霉烯醇的降解效率,在获得ZHD101 晶体结构的基础上,Xu 等进一步研究了ZHD101 分别与底物玉米赤霉烯酮、玉米赤霉烯纯间的复合体结构,通过二者结构的比对,设计了系列突变位点修饰底物结合位点的结构,突变体V153H 对α-玉米赤霉烯醇的催化活性较野生型提高了3.7 倍,但同时保持了对玉米赤霉烯酮的催化活性。
真菌毒素污染给粮食与畜牧业的发展带来了巨大的经济损失,严重威胁着人类的健康和社会的发展。酶法降解真菌毒素技术与传统的物理、化学好微生物脱毒方法相比,具有不可比拟的优势。但目前关于真菌毒素的酶降解技术大部分仍停留在起步阶段,新酶的发掘工作仍举步维艰,在过去的10年中,公开报道的可降解真菌毒素的新酶仅有7个。酶法脱毒技术的应用尚存在以下几方面的问题有待深入探索与研究:
①虽然目前报道的可用于降解真菌毒素的微生物很多,但众多的研究报道中毒素的代谢机理仍缺乏深入研究,制约了新酶的发掘;
②某些真菌毒素的降解依赖于几个酶的协同作用,如在脱羧酶和转氨酶的共同作用下伏马毒素被降解完全,这必将增加新酶发掘的难度;
③同一样品中几种毒素往往共同存在,故在利用酶法降解这些毒素时往往需要多个酶的参与,增加了工艺调控的难度;
④一些酶对真菌毒素的作用是将其降解为毒性较小的化合物,如内酯水解酶降解玉米赤霉烯酮的产物为低毒性的玉米赤霉烯醇,一定程度上***了该酶在毒素降解中的应用。
近年来,生物信息学的发展为新酶的发现提供了更为丰富的技术手段。利用宏基因组技术,通过提取特定环境中的微生物基因组DNA构建基因组文库,从文库中筛选新的功能基因,已成为功能基因筛选的**方法;由于测序技术的发展,已有海量的基因数据资源被公布,根据催化反应的需求,借助计算机辅助筛选,可以从庞大的基因数据库中**获得具有优良催化性能的新型生物酶。而近几年发展起来的无细胞蛋白表达等技术,为酶的高通量筛选提供了有效手段,能够极大地缩短新酶的发掘周期。因此,虽然生物酶法去除真菌毒素的研究起步较晚,发展相对缓慢,但随着相关生物技术的蓬勃发展,相信在不久的将来,越来越多可降解真菌毒素的生物酶将被发掘,并逐步从实验室研究走向实际生产过程。
河南新汉博生物科技有限公司自主研发的新型霉菌毒素酶解产品,将具有降解黄曲霉毒素、玉米赤霉烯酮、呕吐毒素、T-2毒素能力的菌株分别进行固态发酵,之后对分泌的毒素降解酶进行低温干燥处理和优化组合,**终得到对以上四种毒素均具有显著降解能力的产品组合——霉力克。该产品一上市就获得广大用户的认可,数据表明,霉力克对黄曲霉毒素B1的降解率达到89.7%,对玉米赤霉烯酮的降解率达到92.5%,对T-2毒素的降解率达到70.6,对呕吐毒素的降解率达到98.2%,真正从根本上解决困扰养殖界的霉菌毒素问题。
![]() 咨询热线
咨询热线
0373-867-7288
手机:13603711454
地址:河南省长垣县赵堤镇工业园区(尚寨村)